Doppia concimazione nelle piante
La doppia fertilizzazione nelle piante ha un grande significato biologico. Fu scoperto da Navashin nel 1898. Successivamente, diamo un'occhiata più da vicino a come avviene la doppia fecondazione nelle piante. 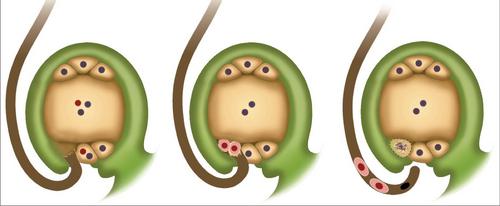
Significato biologico
Il processo di doppia fertilizzazione promuove lo sviluppo attivo del tessuto nutriente. A questo proposito, l'ovulo non immagazzina materiale per il futuro. Questo, a sua volta, spiega il suo rapido sviluppo.
Schema di doppia concimazione
In breve, il fenomeno può essere descritto come segue. La doppia fertilizzazione nelle angiosperme consiste nella penetrazione di due spermatozoi nell'ovaio. Uno si fonde con l'uovo. Ciò contribuisce all'inizio dello sviluppo di un embrione diploide. Il secondo spermatozoo si connette alla cellula centrale. Di conseguenza, si forma un elemento triploide. L'endosperma appare da questa cellula. È un materiale nutriente per l'embrione in via di sviluppo.
Ad
Sviluppo del tubo pollinico
La doppia fecondazione delle angiosperme inizia dopo la formazione di una generazione aploide altamente ridotta. È rappresentato dai gametofiti. La doppia concimazione delle piante da fiore favorisce la germinazione del polline. Inizia con il rigonfiamento del grano e la successiva formazione di un tubo pollinico. Sfonda lo sporoderma nella sua sezione più sottile. Si chiama apertura. Sostanze specifiche vengono emesse dalla punta del tubo pollinico. Ammorbidiscono i tessuti della colonna e lo stigma. A causa di ciò, includono un tubo pollinico. Mentre si sviluppa e cresce, sia gli spermatozoi che il nucleo dalla cellula vegetativa vi entrano. Nella stragrande maggioranza dei casi, il tubo pollinico penetra nel nucleo (megasporangia) attraverso il micropilo dell'ovulo. Questo è raramente fatto in un altro modo. Dopo la penetrazione nel sacchetto germinale, il tubo pollinico si rompe. Di conseguenza, tutto il suo contenuto viene versato all'interno. La doppia fertilizzazione delle piante in fiore continua con la formazione di uno zigote diploide. Questo contribuisce al primo sperma. Il secondo elemento è collegato al nucleo secondario, che si trova nella parte centrale del sacco embrionale. Il nucleo triploide formato viene successivamente trasformato in endosperma. 
Formazione cellulare: informazioni generali
Il processo di doppia fertilizzazione delle piante da fiore viene effettuato da speciali cellule germinali. La loro formazione avviene in due fasi. Il primo stadio è chiamato sporogenesi, il secondo è l'ematogenesi. Nel caso della formazione di cellule maschili, questi stadi sono indicati come microsporogenesi e microematometatogenesi. Nella formazione di elementi genitali femminili, il prefisso cambia in "mega" (o "macro"). La sporogenesi è basata sulla meiosi. Questo è il processo di formazione di elementi aploidi. La meiosi, così come tra i rappresentanti della fauna, è preceduta dalla moltiplicazione delle cellule attraverso divisioni mitotiche.
Ad
Produzione di sperma
La formazione primaria degli elementi genitali maschili viene effettuata in un tessuto speciale di antera. Si chiama archespariale. In esso, come risultato delle mitosi, si verifica la formazione di numerosi elementi, le cellule del polline madre. Quindi entrano nella meiosi. A causa di due divisioni meiotiche, si formano 4 microspore aploidi. Per qualche tempo si trovano nelle vicinanze, formando tetradi. Dopo questo, si disintegrano in grani di polline - microspores individuali. Ognuno degli elementi formati inizia ad essere coperto con due gusci: l'esterno (esino) e l'interno (intin). Quindi inizia la fase successiva: microgametogenesi. A sua volta, consiste di due divisioni successive mitotiche. Dopo la formazione delle prime due cellule: generativa e vegetativa. Successivamente, il primo supera un'altra divisione. Di conseguenza, si formano due cellule maschili: le cellule spermatiche. 
Macroporogenesi e megasporogenesi
Nei tessuti degli ovuli, uno o più elementi archesporiali iniziano a separarsi. Iniziano a crescere rapidamente. A causa di questa attività, diventano significativamente più grandi rispetto al resto delle cellule che li circondano nell'ovulo. Ogni elemento arpore è soggetto a divisione per mitosi una, due o più volte. In alcuni casi, la cellula può immediatamente trasformarsi in madre. La meiosi si verifica al suo interno. Di conseguenza, si formano 4 cellule aploidi. Di regola, il più grande di loro inizia a svilupparsi, trasformandosi in un sacchetto germinale. I tre rimanenti gradualmente degenerano. A questo punto, la macrosporogenesi è completata, inizia l'ematogenesi macro. Durante questo periodo si verificano divisioni mitotiche (la maggior parte delle angiosperme ne ha tre). La citochinesi non accompagna le mitosi. Come risultato di tre divisioni, si forma un sacco embrionale con otto nuclei. Successivamente si isolano in cellule indipendenti. Questi elementi sono distribuiti in un certo modo dalla borsa germinale. Una delle cellule isolate, che in realtà è una cellula uovo, insieme ad altre due sinergidi, ha luogo al micropilo, in cui avviene la penetrazione dello sperma. In questo processo, i sinergisti svolgono un ruolo molto significativo. Contengono enzimi che contribuiscono alla dissoluzione delle membrane sui tubi di polline. Nel lato opposto del sacco germinale si trovano le altre tre cellule. Si chiamano antipodi. Con l'aiuto di questi elementi, i nutrienti vengono trasferiti dagli ovuli al sacco germinale. Le restanti due celle si trovano nella parte centrale. Spesso si uniscono. Come risultato della loro connessione, si forma una cellula diploide centrale. Dopo la doppia fecondazione e lo sperma penetra nell'ovaia, uno di essi, come detto sopra, si fonderà con la cellula uovo. 
Dispone di tubo pollinico
La doppia fertilizzazione è accompagnata dalla sua interazione con i tessuti sporofiti. È abbastanza specifico. Questo processo è governato dall'attività dei composti chimici. Si scopre che se il polline viene lavato acqua distillata perderà la capacità di germinare. Se la soluzione risultante viene concentrata e quindi elaborata, verrà nuovamente completata. Lo sviluppo del tubo pollinico dopo la germinazione controlla il tessuto del pistillo. Ad esempio, nel cotone, la sua crescita nell'uovo dura circa 12-18 ore. Tuttavia, già dopo 6 ore è abbastanza possibile determinare a quale ovulo andrà il tubo pollinico. Questo è comprensibile perché inizia la distruzione della sinergia. Allo stato attuale, non è stabilito in che modo la pianta possa dirigere lo sviluppo del tubo nella giusta direzione e in che modo il sinergizzante apprende l'approssimazione. 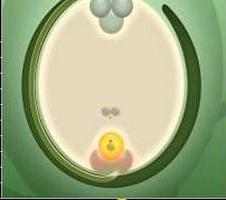
"Divieto" sul selfing
È spesso osservato nelle piante da fiore. Questo fenomeno ha le sue caratteristiche. Il "divieto" di autoimpollinazione si manifesta nel fatto che lo sporofito "identifica" il proprio ematofito maschile e non gli consente di partecipare alla fecondazione. Allo stesso tempo, in alcuni casi, la germinazione del proprio polline non si verifica sullo stigma del pistillo. Tuttavia, di regola, la crescita del tubo inizia ancora, ma successivamente si interrompe. Di conseguenza, il polline non raggiunge l'uovo e, di conseguenza, non si verifica una doppia concimazione. Darwin è stato notato questo fenomeno. Quindi, ha trovato in Primula fiori primaverili due forme. Alcuni di essi erano lunghi steli con stami corti. Altri sono a colonna breve. In essi i filamenti staminati erano lunghi. Le piante a stelo corto si differenziano per il grande polline (il doppio rispetto agli altri). Allo stesso tempo, le cellule nelle papille dello stigma sono piccole. Questi segni sono controllati da un gruppo di geni strettamente intrecciati. 
I recettori
La doppia fertilizzazione è efficace quando il polline viene trasferito da una forma all'altra. Per il riconoscimento dei propri elementi sono responsabili molecole recettore speciali. Sono composti complessi di carboidrati con proteine. È stato stabilito che forme di cavolo selvatico, che non producono queste molecole recettoriali nei tessuti dello stigma, possono auto-impollinarsi. Per le piante normali, i composti di carboidrati e proteine compaiono il giorno prima dell'apertura del fiore. Se apri un germoglio e lo processi con il tuo polline due giorni prima della sua fioritura, allora si verificherà una doppia fecondazione. Se questo è fatto il giorno prima dell'apertura, non lo sarà.
Ad
alleli
È interessante notare che in alcuni casi l'"autoincompatibilità" del polline nelle piante è stabilita da una serie di più elementi di un singolo gene. Questo fenomeno è simile all'incompatibilità del trapianto di tessuto negli animali. Tali alleli sono indicati con la lettera S. Il numero nella popolazione di questi elementi può raggiungere decine o anche centinaia. Ad esempio, se il genotipo della pianta produttrice di uova è s1s2, e s2s3 che produce polline, durante l'impollinazione incrociata, la germinazione sarà annotata solo nel 50% delle particelle di polvere. Questi saranno quelli che portano l'allele S3. Se ci sono diverse dozzine di elementi, la maggior parte del polline germinerà normalmente durante l'impollinazione incrociata, mentre l'autoimpollinazione è completamente prevenuta. 
In conclusione
In contrasto con le gimnosperme, che sono caratterizzate dallo sviluppo di un endosperma aploide sufficientemente potente, indipendentemente dalla fertilizzazione, nelle angiosperme, il tessuto si forma solo in questo singolo caso. Dato l'enorme numero di generazioni, ha conseguito un notevole risparmio energetico. A quanto pare, l'aumento del grado di ploidia dell'endosperma contribuisce a una crescita tissutale più rapida rispetto agli strati diploidi dello sporofito.