Complesso di Golgi: descrizione
Il complesso Golgi fu scoperto nel 1898. Questa struttura a membrana è progettata per eliminare i composti che sono sintetizzati nel reticolo endoplasmatico. Inoltre faremo conoscere questo sistema. 
Complesso del Golgi: costruzione
Il dispositivo è una pila di serbatoi a forma di disco a membrana. Queste buste sono un po 'estese verso i bordi. Il sistema di bolle Golgi è associato ai serbatoi. Nelle cellule animali, sono presenti una grande o più pile, che sono collegate da tubi, in cellule vegetali Dictyosomes sono rilevati (diversi stack separati). Il complesso Golgi comprende tre dipartimenti. Sono circondati da bolle di membrana:
- cis-più vicino al nucleo;
- mediale;
- dipartimento trance (il più lontano dal nucleo).
Questi sistemi sono contraddistinti da un set di enzimi. Nel dipartimento cis, la prima sacca viene definita "il serbatoio di salvataggio". Con il suo aiuto, i recettori che provengono dalla rete intermedia endoplasmatica tornano indietro. Dipartimento di enzimi cis chiamato fosfoglicosidasi. Aggiunge fosfato al mannosio (carboidrati). Nella parte mediale si trovano due enzimi. Questo, in particolare, la mennadiasi e la N-acetilglucosamina transferasi. Quest'ultimo aggiunge glicosammine. Enzimi del transpartimento: peptidasi (esegue proteolisi) e transferasi (con il suo aiuto avviene il trasferimento di gruppi chimici). 
Complesso di Golgi: funzioni
Questa struttura prevede la separazione delle proteine nei seguenti tre flussi:
- Lisosomiale. Secondo esso, le proteine glicate penetrano nel dipartimento cis dell'apparato di Golgi. Alcuni di loro sono fosforilati. Di conseguenza, il mannosio-6-fosfato si forma - enzimi mercososomici. In futuro, queste proteine fosfatate entreranno nei lisosomi e non saranno modificate.
- Esocitosi costitutiva (secrezione). Questo flusso include proteine e lipidi, che sono diventati componenti dell'apparato di superficie cellulare, incluso il glicocalice. Inoltre, possono essere presenti composti che fanno parte della matrice extracellulare.
- Secrezione indotta Le proteine, che funzionano al di fuori della cellula, apparati di superficie, penetrano in questo flusso nell'ambiente interno del corpo. La secrezione indotta è caratteristica delle cellule secretorie.
Il complesso del Golgi è coinvolto nella formazione della secrezione mucosa - mucopolisaccaridi (glicosaminoglicani). Il dispositivo forma anche i componenti di carboidrati del glicocalice. Sono per lo più glicolipidi. Il sistema prevede anche la solfatazione di elementi proteici e carboidrati. Il complesso di Golgi è coinvolto nella parziale proteolisi delle proteine. In alcuni casi, a causa di ciò, il composto dall'inattivo entra nella forma attiva (ad esempio, la proinsulina viene trasformata in insulina). 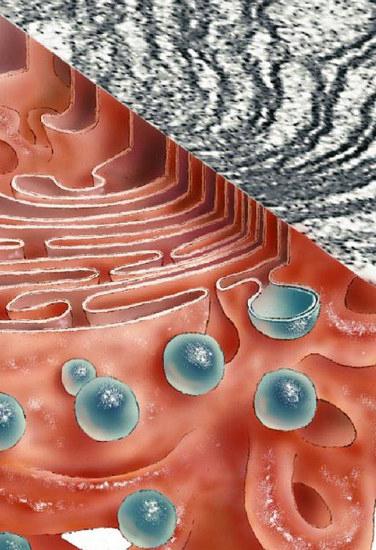
Composti in movimento dal reticolo endoplasmatico (EPS)
Il complesso è asimmetrico. Situato più vicino al nucleo della cellula includono le proteine più immature. Vesicles, vescicole di membrana, sono continuamente attaccati a queste sacche. Derivano dal reticolo granulare endoplasmatico. Sulle sue membrane passa il processo di sintesi proteica da parte dei ribosomi. Il trasporto di composti dal reticolo endoplasmatico al complesso di Golgi è indiscriminato. Allo stesso tempo, le proteine piegate in modo errato o incompleto continuano a rimanere nell'EPS. Il movimento inverso dei composti nel reticolo endoplasmatico richiede una speciale sequenza di segnali ed è reso possibile dal legame di queste sostanze ai recettori di membrana nel compartimento cis.
Ad
Modifica delle proteine
Nei serbatoi della maturazione complessa si verificano composti che sono destinati alla secrezione, transmembrana, lisosomiale e altre sostanze. Queste proteine si spostano successivamente lungo i serbatoi verso gli organelli. Le loro modificazioni cominciano in loro - fosfatazione e glicosilazione. Durante il primo processo, un residuo è collegato alle proteine. acido fosforico. Nella glicosilazione, gli zuccheri complessi sono ancorati attraverso l'atomo di ossigeno. Diversi serbatoi contengono vari enzimi catalitici. Di conseguenza, i processi successivi differenti avvengono con la maturazione delle proteine in essi. Indubbiamente, un tale fenomeno graduale dovrebbe essere controllato. Come una sorta di residui di polisaccaride "di qualità" (mannose, principalmente) vengono utilizzati. Etichettano le proteine in scadenza. Ulteriore movimento attraverso i serbatoi di composti non è completamente compreso dalla scienza, nonostante il fatto che le sostanze resistenti rimangano meno o più associate a una borsa. 
Trasporto di proteine dall'apparecchio
Dal complesso trans-dipartimentale nascono bolle. Contengono composti proteici completamente maturi. La funzione principale del complesso è lo smistamento delle proteine che lo attraversano. Nell'apparecchio, la formazione di un "flusso proteico tridirezionale" - maturazione e trasporto:
Ad
- composti membrana plasmatica.
- Segreti.
- Enzimi lisosomiali
Attraverso il trasporto vescicolare, le proteine che sono passate attraverso il complesso di Golgi vengono consegnate a determinati siti in conformità con i "tag". Anche questo processo non è completamente compreso dalla scienza. È stato stabilito che il trasporto di proteine dal complesso richiede la partecipazione di specifici recettori di membrana. Riconoscono il composto e forniscono l'attracco selettivo della vescicola e l'uno o l'altro organello. 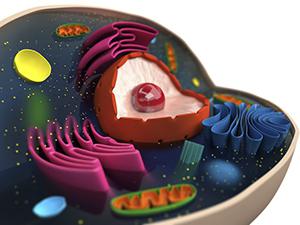
Formazione del lisosoma
Attraverso l'apparato passa molti enzimi idrolitici. L'aggiunta dell'etichetta sopra menzionata viene effettuata con la partecipazione di due enzimi. Il riconoscimento specifico delle idrolasi lisosomiali da parte degli elementi della loro struttura terziaria e l'aggiunta di N-acetilglucosamina fosfato è effettuata dalla N-acetilglucosamina fosfotransferasi. Il fosfoglicoside, il secondo enzima, rimuove la N-acetilglucosamina, determinando la formazione di un'etichetta M6F. A sua volta, è riconosciuto dalla proteina recettore. Con il suo aiuto, le idrolasi vengono consegnate alle vescicole e ai loro mandati nei lisosomi. In condizioni acide, il fosfato viene scisso dall'idrolasi matura in esse. Se ci sono disturbi nell'attività della N-acetilglucosammina fosfotransferasi dovuta a mutazioni oa causa di difetti genetici nel recettore M6F, tutti gli enzimi lisosomiali vengono consegnati di default alla membrana esterna. Vengono quindi secreti in condizioni extracellulari. È stato anche stabilito che alcuni dei recettori M6P sono anche trasportati sulla membrana esterna. Eseguono il ritorno degli enzimi lisosomiali casualmente catturati dall'ambiente esterno all'interno della cellula durante l'endocitosi. 
Trasporto di sostanze sulla membrana esterna
Solitamente, anche allo stadio della sintesi, i composti proteici della membrana esterna con le loro regioni idrofobiche sono incorporati nella parete del reticolo endoplasmatico. Quindi vengono consegnati al complesso di Golgi. Da lì vengono trasportati sulla superficie della cella. Nel processo di fusione del plasmalemma e delle vescicole, tali composti non vengono rilasciati nell'ambiente.
secrezione
Quasi tutti i composti prodotti nella cellula (sia di natura proteica che non proteica) passano attraverso il complesso di Golgi. Lì si sviluppano in vescicole secretorie. Nelle piante con dictyo, il materiale viene quindi prodotto. parete cellulare.